하플로그룹 M (mtDNA)
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
하플로그룹 M (mtDNA)은 미토콘드리아 DNA(mtDNA)의 하플로그룹으로, 인류의 모계 유전 계통을 나타낸다. M은 자매 하플로그룹인 N과 함께 아프리카를 벗어난 주요 계통이며, 아프리카 외부에서 발견되는 모든 토착 계통은 M 또는 N에 속한다. M의 기원에 대해서는 아프리카 기원설과 아시아 기원설이 있으며, 아프리카 기원설은 M1을 포함한 아프리카 내의 M 하위 그룹에 근거하고, 아시아 기원설은 아시아, 특히 인도, 중국, 일본, 한국에서 M의 높은 빈도와 다양한 하위 계통에 근거한다. M은 아시아에서 가장 흔하며, 하위 계통은 전 세계적으로 분포하며 특정 지역에서 높은 빈도를 보인다.

더 읽어볼만한 페이지
- 인류 미토콘드리아 DNA 하플로그룹 - 하플로그룹 L3 (mtDNA)
하플로그룹 L3 (mtDNA)는 6만 년에서 8만 년 전에 발생하여 호모 사피엔스의 출아프리카 대이주 시기에 기원한 미토콘드리아 DNA 하플로그룹으로, 아프리카 외부 인구의 주류를 형성한 하플로그룹 M과 N의 기원이며 인류 이동 경로 연구에 활용되고 있다. - 인류 미토콘드리아 DNA 하플로그룹 - 미토콘드리아 이브
미토콘드리아 이브는 현생 인류의 미토콘드리아 DNA 분석을 통해 추정되는, 모계 혈통으로만 유전되는 DNA를 가진 모든 인류의 가장 가까운 공통 여성 조상으로, 약 10만 년에서 20만 년 전 아프리카에 살았던 것으로 추정되지만, 최초의 여성이거나 유일한 생존 여성은 아니다.
2. 기원
하플로그룹 M과 그 자매 하플로그룹 N의 지리적 기원에 대해서는 논쟁이 있다. 두 계통 모두 아프리카 이주에 관여했던 주요 생존 계통이었던 것으로 여겨지는데, 그 이유는 아프리카 외부에서 발견되는 모든 토착 계통이 하플로그룹 M 또는 하플로그룹 N에 속하기 때문이다. 과학자들은 하플로그룹 M과 N을 정의하는 돌연변이가 아프리카를 떠나기 전 아프리카에서 발생했는지, 아니면 아프리카를 떠난 후 아시아에서 발생했는지 확신하지 못하고 있다. 하플로그룹 M의 기원을 밝히는 작업은 M1을 가진 사람들의 초기 역이주(아시아에서 아프리카로)로 인해 더욱 복잡해진다.[10]
절대적인 연도로 표현한 기원 시기는 재구성에서 남아시아와 동아시아의 M의 연령에 대한 서로 다른(하지만 겹치는) 범위가 제시되었기 때문에 매우 불확실하게 알려져 있다. 하플로그룹 M은 L3 이후 약 1만 년 또는 최대 2만 년 후, 최근 아프리카 이주 사건과 비슷하거나 약간 후에 출현했을 것으로 추정된다.
M1은 가장 먼저 분기되어 아프리카를 떠나지 않은 계통으로 아프로-아시아어족과 관련이 있는 것으로 알려져 있다.
2. 1. 아프리카 기원설
하플로그룹 M과 그 자매 하플로그룹 N의 지리적 기원에 대해서는 논쟁이 있다. 두 계통 모두 아프리카 이주에 관여했던 주요 생존 계통이었던 것으로 여겨지는데, 그 이유는 아프리카 외부에서 발견되는 모든 토착 계통이 하플로그룹 M 또는 하플로그룹 N에 속하기 때문이다. 과학자들은 하플로그룹 M과 N을 정의하는 돌연변이가 아프리카를 떠나기 전 아프리카에서 발생했는지, 아니면 아프리카를 떠난 후 아시아에서 발생했는지 확신하지 못하고 있다. 하플로그룹 M의 기원을 밝히는 작업은 M1을 가진 사람들의 초기 역이주(아시아에서 아프리카로)로 인해 더욱 복잡해진다.[10]하플로그룹 M의 기원에 대한 많은 논의는 아프리카에서 발견되는 거대 하플로그룹 M의 유일한 변이체인 하플로그룹 M1과 관련이 있다.[35] 아프리카에서 M1이 존재하는 것에 대한 두 가지 가능성이 잠재적인 설명으로 고려되었다.
# M은 나중에 아프리카의 M1과 유라시아에서 더 일반적으로 발견되는 M을 모두 낳은 고대 인구에 존재했다.[12]
# 아프리카에서 M1이 존재하는 것은 아프리카 이주 후 어느 시점에 일어난 아시아로부터의 역이주 결과이다.[10]
2009년, 두 개의 독립적인 연구에서 마다가스카르에서 발견된 희귀하고 깊이 뿌리내린 M 하플로그룹의 하위 분류인 M23을 보고했다.[13][14] M23의 기원은 아직 명확하지 않으며, 이전에는 서유라시아 기원이 제시되었다. M23 하플로그룹은 고대이며 마다가스카르 전역에서 발견된다. 상당한 아프리카 구성 요소를 가진 아시아 구성 요소로 추정된다. M23은 두바이에서 단 한 명의 개인에게서만 발견되었다.[15]
2. 2. 아시아 기원설
하플로그룹 M과 그 자매 하플로그룹 N의 지리적 기원에 대해서는 논쟁이 있다. 두 계통은 모두 아프리카 이주에 관여했던 주요 생존 계통으로 여겨지는데, 아프리카 외부에서 발견되는 모든 토착 계통이 하플로그룹 M 또는 N에 속하기 때문이다. 과학자들은 하플로그룹 M과 N을 정의하는 돌연변이가 아프리카에서 발생했는지, 아니면 아시아에서 발생했는지 확신하지 못하고 있다. 하플로그룹 M의 기원을 밝히는 작업은 M1을 가진 사람들의 초기 역이주(아시아에서 아프리카로)로 인해 더욱 복잡해진다.[10]절대적인 연도로 표현한 기원 시기는 재구성에서 남아시아와 동아시아의 M의 연령에 대한 서로 다른(하지만 겹치는) 범위가 제시되었기 때문에 매우 불확실하게 알려져 있다. 하플로그룹 M은 L3 이후 약 1만 년 또는 최대 2만 년 후, 최근 아프리카 이주 사건과 비슷하거나 약간 후에 출현했을 것이다. 2009년에는 마다가스카르에서 발견된 희귀하고 깊이 뿌리내린 M 하플로그룹의 하위 분류인 M23이 보고되었다.[13][14]
M23의 기원은 아직 명확하지 않으며, 이전에는 서유라시아 기원이 제시되었다. M23 하플로그룹은 고대이며 마다가스카르 전역에서 발견된다. 상당한 아프리카 구성 요소를 가진 아시아 구성 요소로 추정된다. M23은 두바이에서 단 한 명의 개인에게서만 발견되었다.[15]
이 이론에 따르면, 초기 하플로그룹 L3 계통을 가진 해부학적으로 현생 인류가 아프리카 동부에서 아시아로 이주했다. 아시아 어딘가에서 초기 L3 계통은 하플로그룹 M과 N을 낳았다. 그 후 초기 L3 계통은 아프리카 외부에서 빈도가 낮기 때문에 유전적 부동에 의해 사라졌다. 아시아가 거대 하플로그룹 M의 기원이라는 가설은 다음과 같은 사실들로 뒷받침된다.
- 전 세계적으로 거대 하플로그룹 M의 가장 높은 빈도는 아시아, 특히 인도 아대륙, 중국, 일본, 한국에서 관찰되는데, 그 빈도는 60~80%에 이른다.[16][17][18]
- 인도 서부, 중부, 남부, 동부의 하플로그룹 M2, M38, M54, M58, M33, M6, M61, M62의 깊은 시간 깊이(5만 년 이상)와 거대 하플로그룹 M의 분포는 거대 하플로그룹 M이 인도 집단에서 발생했을 가능성을 배제하지 않는다.[19]
- 아프리카 특이적 M1을 제외하고 인도에는 하플로그룹 M의 근원에서 직접 유래된 여러 M 계통이 있다.[16][18]
- 하플로그룹 M의 하위 계통 중 아프리카에서 발견되는 것은 M1과 M23 두 가지뿐이며, 수많은 하위 계통이 아프리카 외부에서 발견된다.[16][10]
M1에 관한 구체적인 내용은 다음과 같다.
- 하플로그룹 M1은 아프리카에서 지리적 분포가 제한적이며, 주로 북아프리카와 동아프리카에서 낮거나 중간 정도의 빈도로 발견된다. M이 아프리카 이주 전후에 아프리카에서 기원했다면, 더 넓은 분포를 보일 것으로 예상된다.[18]
- Gonzalez 등(2007)에 따르면, M1은 비교적 최근에 확장된 것으로 보인다. 이 연구에서 M1은 아시아 독점 M 계통보다 더 젊은 합류 연령을 가졌다.[10]
- 아프리카에서 M1의 지리적 분포는 주로 북아프리카/적도 상부[10]이며, 대부분 아프로아시아어족 화자에게 국한되어 있는데,[20] 이는 유사한 시간 깊이를 가진 하플로그룹 L3 및 L2의 하위 계통의 사하라 이남 분포와 일치하지 않는다.[35]
- M1 계통의 기저 계통 중 하나는 북서아프리카와 근동에서 발견되었지만 동아프리카에는 없다.[10]
- M1은 아프리카에 국한되지 않는다. 지중해 지역에서 비교적 흔하며, 이베리아에서 정점을 이룬다. M1은 또한 아라비아 반도 남부에서 아나톨리아까지, 레반트에서 이란까지 중동에 널리 분포되어 있다. 또한 M1 하플로타입은 카프카스와 트랜스카프카스에서 간헐적으로 관찰되었으며, L 계통은 동반되지 않았다.[10][35] M1은 중앙아시아에서도 발견되었으며, 티베트까지 이른 것으로 보인다.[10]
- 거대 하플로그룹 M의 M1 하위 계통의 합류 연령이 하플로그룹 U6 (아프리카에 존재하는 것은 서아시아에서의 역이주 때문인 유라시아 하플로그룹)과 일치하고, 아프리카에서 U6의 분포가 M1과 동일한 북아프리카와 뿔의 아프리카 인구에 제한되어 있다는 사실은 M1과 U6가 아시아에서 아프리카로의 동일한 인구 확장의 일부였을 것이라는 시나리오를 뒷받침한다.[20]
- 서아시아에서 아프리카로의 M1과 U6를 가진 사람들의 이주(4만~4만 5천 년 전)에 대한 제안된 시기는 북아프리카의 사막 지역을 감소시켜 레반트에서의 진입을 용이하게 한 기후 조건의 변화와 일치한다는 사실로도 뒷받침된다. 이러한 기후 변화는 또한 하플로그룹 U5(하플로그룹 U6의 유럽 자매 계통)를 가진 인구에 의한 유럽 거주와 시간적으로 일치한다.[20]
3. 확산
여러 연구에서 현대 하플로그룹 M의 조상들은 아프리카에서 아시아 해안 지역을 따라 아프리카의 뿔을 지나 남쪽 경로를 통해 뉴기니와 오스트레일리아까지 이동했을 것이라고 제안한다.[23][24] 최근 연구들은 아프리카에서 단 한 번의 이동이 있었으며 하플로그룹 M과 N이 동일한 이동의 일부였다는 것을 나타낸다.[23][24] 이는 아프리카에서 오스트레일리아까지 해변을 따라 이동하는 경로를 따라 제안된 여러 잔존 집단에 대한 분석을 기반으로 한다.[23][24]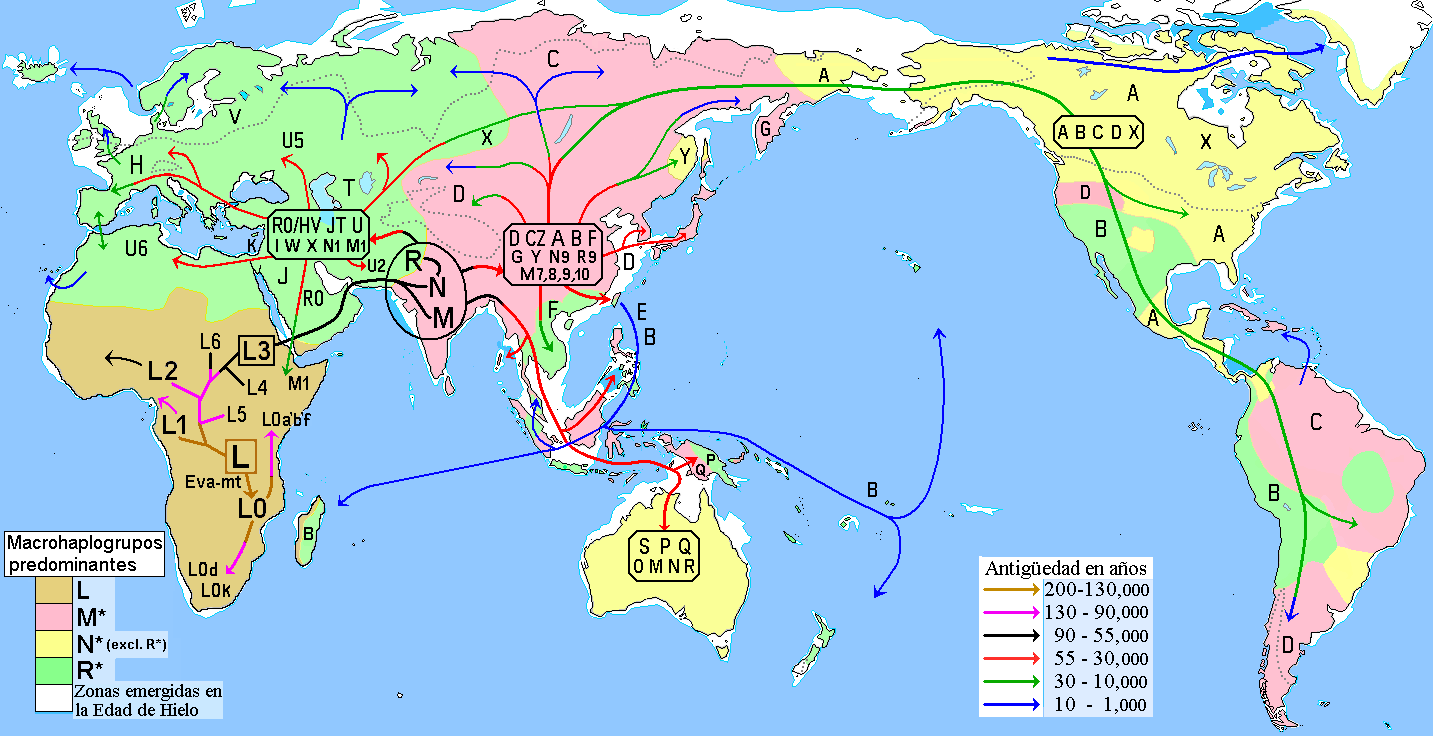
Abu-Amero 등의 2008년 연구는 아라비아 반도가 아프리카 밖으로 나가는 주요 경로였을 수 있다고 제안한다.[25] 그러나 이 지역에는 하플로그룹 M과 N의 토착 분지가 부족하기 때문에, 이 지역이 고대 인구 확장 중심지라기보다는 최근의 인류 이동 수용 지역이었을 것이라고 제안한다.[25]
4. 분포
하플로그룹 M은 아시아에서 가장 흔한 mtDNA 하플로그룹으로, 모든 모계 혈통의 60%를 차지한다.[154] 특히 안다만 제도 원주민과 말레이시아 네그리토 부족에서 거의 100%에 달하는 최고 빈도를 보인다. 일본과 티베트에서는 모계 혈통의 약 70%, 인도와 한국에서는 약 60%를 나타낸다.[156][157] 중국에서는 평균 50% 정도이나, 남부에서는 비교적 적고 북동부로 갈수록 높아진다.[154]
동남아시아에서는 필리핀인의 약 42%[158], 베트남인의 37~48%, 빈투언 성 참족의 32%[154], 라오스인의 43%[154] 정도로 나타난다. 오세아니아에서는 파푸아 뉴기니 제도에서 28~42% 정도이다.
북동아프리카에서는 소말리아인, 리비아인, 오로모인에게서 높게 나타난다.[159] 투아레그 지역에서는 말리와 부르키나파소에서 M1a2를 중심으로 상당한 비율로 나타난다.[160] 유럽에서는 이베리아 반도나 이탈리아 등 지중해 연안에서 비교적 많이 발견된다.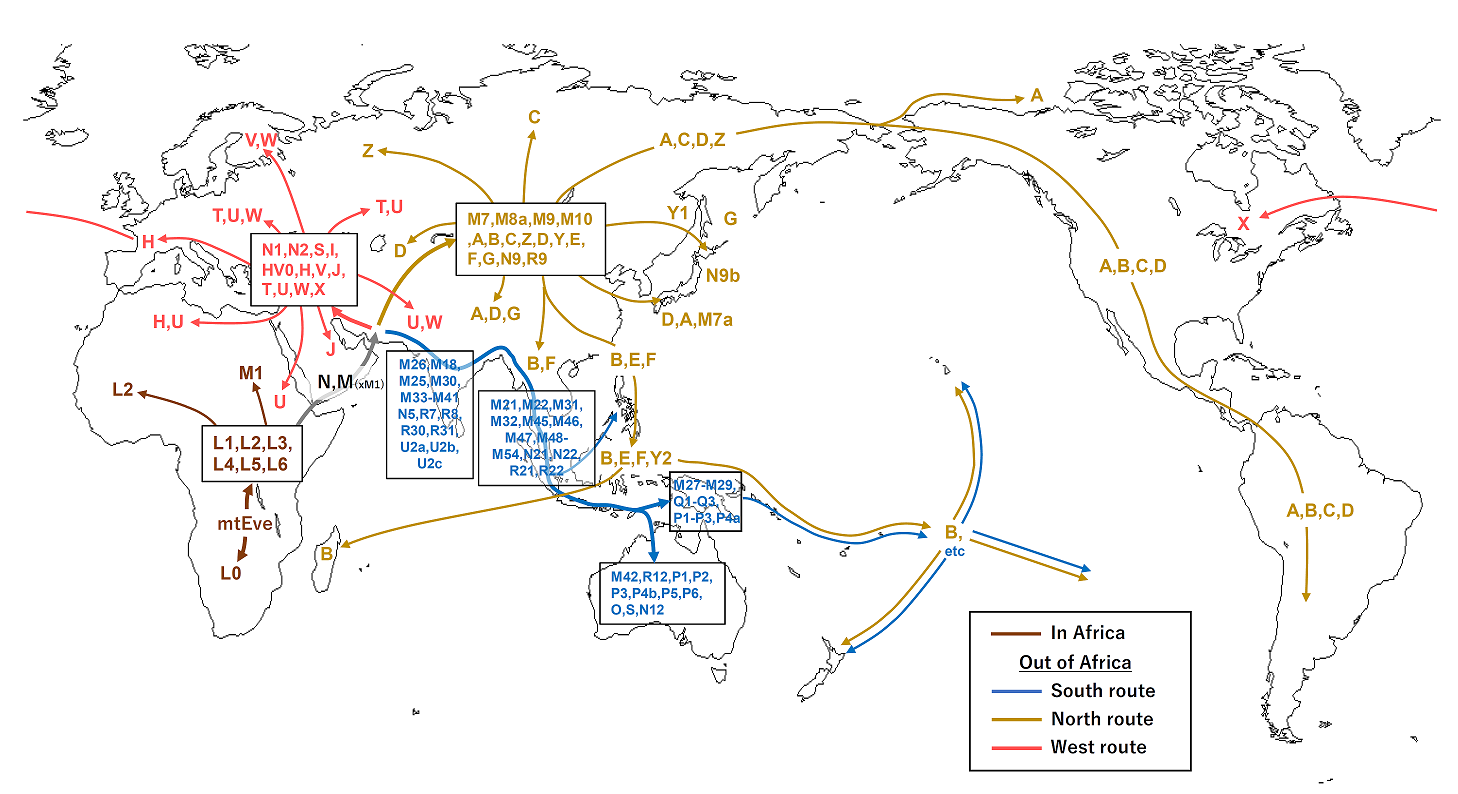
여러 연구에 따르면 현대 하플로그룹 M의 조상들은 아프리카에서 아시아 해안 지역을 따라 아프리카의 뿔을 지나 남쪽 경로를 통해 뉴기니와 오스트레일리아까지 이동했을 것으로 추정된다.
4. 1. 하위 계통
하플로그룹 M은 아시아에서 가장 흔하게 나타나며, 모계 혈통의 60%를 차지한다.[154] 특히 안다만 제도 원주민과 말레이시아 네그리토족에서 매우 높은 빈도를 보인다.[155] 일본과 티베트에서는 모계 혈통의 70%[156], 인도와 한국에서는 60%[156][157], 중국에서는 평균 50%[154] 정도로 나타난다. 동남아시아, 오세아니아, 북동아프리카에서도 비교적 높은 수치를 보인다.하플로그룹 M은 매우 다양한 하위 계통을 가지고 있으며, 주요 하위 계통은 다음과 같다.
- M1: 나일강 유역, 아프리카의 뿔, 마그레브, 사하라, 지중해 연안, 중동 등지에 분포한다.
- M2: 남아시아, 특히 인도 동남부와 방글라데시에서 높은 빈도를 보인다.
- M3: 남아시아, 특히 인도 서북부에서 높은 빈도를 보인다.
- M4'30: 남아시아, 사우디아라비아 동부에 분포한다.
- M5: 남아시아에 분포한다.
- M6: 남아시아에 분포한다.
- M7: 동아시아, 동남아시아에 분포하며, 특히 일본, 남중국, 베트남, 라오스, 태국에서 높은 빈도를 보인다.
- M8: 중국, 태국 (리수족), 인도 등에 분포한다.
- M8a: 동아시아, 북아시아에 분포한다.
- CZ: 태국 (흐몽족)에 분포한다.
- M9: 동아시아, 중앙아시아에 분포하며, 특히 티베트에서 높은 빈도를 보인다.
- E: 특히 타이완 원주민, 동남아시아 해안, 마리아나 제도에서 높은 빈도를 보인다.
- M10: 동아시아, 동남아시아, 중앙아시아, 시베리아, 유럽 동북부에 드물게 분포한다.
- M11: 동아시아에 분포하며, 중국에서 비교적 높은 빈도를 보인다.
- M12'G:
- M12: 중국 하이난(리족)에서 높은 빈도를 보이며, 다른 동아시아, 동남아시아 지역에도 분포한다.
- G: 아시아 여러 지역에 산발적으로 분포하며, 일본, 몽골, 티베트, 캄차카반도 원주민에서 특히 높은 빈도를 보인다.
- M13'46'61:
- M13
- M46
- M61
- M14: 오스트레일리아에 분포한다.
- M15: 오스트레일리아에 분포한다.
- M17: 루손섬, 참족, 몬족, 태국, 라오스 등에 분포한다.
- M21: 동남아시아에 드물게 분포한다.
- M23'75:
- M23: 마다가스카르, 남아프리카에 분포한다.
- M75: 중국에 분포한다.
- M24: 태국, 캄보디아, 중국 등에 분포한다.
- M29'Q:
- M29: 멜라네시아에 분포한다.
- Q: 멜라네시아, 오스트레일리아 원주민에 분포한다.
- M31: 옹게인에 분포한다.
- M35: 네팔, 인도에 분포한다.
- M71: 인도, 동남아시아에 분포한다.
- M74: 태국, 캄보디아, 베트남, 중국, 타이완 등에 분포한다.
- M80'D:
- M80: 팔라완섬에 분포한다.
- D: 아시아, 아메리카 원주민에 분포하며, 유럽에 드물게 분포한다.
분류:하플로그룹 M (mtDNA)
5. 계통 발생
이 글은 만니스 반 오번(Mannis van Oven)과 만프레드 카이저(Manfred Kayser)의 논문 "전 세계 인간 미토콘드리아 DNA 변이의 업데이트된 포괄적 계통 발생 나무"[150] 및 그 이후의 연구 결과를 바탕으로 한 하플로그룹 M의 하위 분류 계통 발생 나무이다.
- '''M'''
- *'''M1'''
- **M1a
M1a1
*M1a1a
*M1a1b
**M1a1b1
*M1a1c
*M1a1d
*M1a1e
*M1a1f
M1a2
*M1a2a
*M1a2b
M1a3
*M1a3a
*M1a3b
M1a4
M1a5
- **M1b
M1b1
*M1b1a
M1b2
*M1b2a
- *'''M2'''
- **M2a
M2a1
M2a2
M2a3
- **M2b
M2b1
M2b2
- *M3
- **M3a
- *M4'45
- **M4
M4a
M4b
*M4b1
- **M18'38
M18
M38
- **M30
M30a
M30b
M30c
*M30c1
**M30c1a
***M30c1a1
M30d
- **M37
M37a
- **M43
- **M45
- *M5
- **M5a
M5a1
*M5a1a
*M5a1b
M5a2
*M5a2a
- *M6
- *'''M7'''
- **M7a
M7a1
*M7a1a
**M7a1a1
***M7a1a1a
**M7a1a2
**M7a1a3
**M7a1a4
***M7a1a4a
**M7a1a5
**M7a1a6
**M7a1a7
*M7a1b
M7a2
*M7a2a
*M7a2b
- **M7b'c'd'e
M7b'd
*M7b
**M7b1'2
***M7b1
***M7b2
M7b2a
M7b2b
M7b2c
**M7b3
***M7b3a
*M7d
M7c'e
*M7c
**M7c1
***M7c1a
***M7c1b
M7c1b1
**M7c2
***M7c2a
**M7c3
***M7c3a
***M7c3b
***M7c3c
*M7e
- *'''M8'''
- **M8a
M8a1
M8a2
*M8a2a
*M8a2b
- **'''CZ'''
'''C'''
'''Z'''
- *'''M9'''
- **M9a'b'c'd
M9a'c'd
*M9a'd
**M9a
***M9a1
***M9a2
***M9a3
**M9d
*M9c
M9b
- **'''E'''
- *M10'42
- **M10
M10a
*M10a1
*M10a2
- **M42
M42a
- *M11
- **M11a
- **M11b
- *M12'G
- **M12
M12a
- **'''G'''
- *M13
- **M13a
M13a1
- *M14
- *M15
- *M21
- **M21a'b
M21a
M21b
- **M21c'd
M21c
M21d
- *M22
- *M23
- *M25
- *M27
- **M27a
- **M27b
- **M27c
- *M28
- **M28a
- **M28b
- *M29'Q
- **M29
M29a
M29b
- **'''Q'''
- *M31
- **M31a
M31a1
*M31a1a
*M31a1b
M31a2
- **M31b'c
M31b
*M31b1
*M31b2
M31c
- *M32'56
- **M32
M32a
M32c
- **M56
- *M33
- **M33a
- **M33b
- **M33c
- *M34
- **M34a
- *M35
- **M35a
- **M35b
- *M36
- **M36a
- *M39
- **M39a
- *M40
- **M40a
- *M41
- *M44'52
- **M44
- **M52
- *M46
- *M47'50
- **M47
- **M50
- *M48
- *M49
- *M51
- *M52’58
- *M52
- *'''D'''
참조
[1]
논문
The Expansion of mtDNA Haplogroup L3 within and out of Africa
2012-03
[2]
논문
Ancestral mitochondrial N lineage from the Neolithic 'green' Sahara
2019-03
[3]
논문
Most of the extant mtDNA boundaries in South and Southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans
2004
[4]
논문
Ancestral mitochondrial N lineage from the Neolithic 'green' Sahara
2019-03
[5]
논문
Carriers of human mitochondrial DNA macrohaplogroup M colonized India from southeastern Asia
2016
[6]
논문
Where West Meets East: The Complex mtDNA Landscape of the Southwest and Central Asian Corridor
1999
[7]
논문
Updating Phylogeny of Mitochondrial DNA Macrohaplogroup M in India: Dispersal of Modern Human in South Asian Corridor
2009
[8]
논문
Mitochondrial HVRI and whole mitogenome sequence variations portray similar scenarios on the genetic structure and ancestry of northeast Africans
2021-02
[9]
논문
Carriers of Mitochondrial DNA Macrohaplogroup N Lineages Reached Australia around 50,000 Years Ago following a Northern Asian Route
2015
[10]
논문
Mitochondrial lineage M1 traces an early human backflow to Africa
2007
[11]
논문
Correcting for Purifying Selection: An Improved Human Mitochondrial Molecular Clock
2009-06
[12]
논문
Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern Africa
1999-12
[13]
논문
Complete mitochondrial sequences for haplogroups M23 and M46: insights into the Asian ancestry of the Malagasy population
2009
[14]
논문
A new deep branch of eurasian mtDNA macrohaplogroup M reveals additional complexity regarding the settlement of Madagascar
2009
[15]
논문
Tracing Arab-Islamic Inheritance in Madagascar: Study of the Y-chromosome and Mitochondrial DNA in the Antemoro
2013-11-22
[16]
논문
Phylogeny and antiquity of M macrohaplogroup inferred from complete mt DNA sequence of Indian specific lineages
2005
[17]
논문
Sequence polymorphisms of the mitochondrial DNA control region and phylogenetic analysis of mtDNA lineages in the Japanese population
2003
[18]
논문
In situ origin of deep rooting lineages of mitochondrial Macrohaplogroup 'M' in India
2006-12
[19]
논문
Updating Phylogeny of Mitochondrial DNA Macrohaplogroup M in India: Dispersal of Modern Human in South Asian Corridor
2009
[20]
논문
The mtDNA Legacy of the Levantine Early Upper Palaeolithic in Africa
2006-12-15
[21]
논문
The Genetic Heritage of the Earliest Settlers Persists Both in Indian Tribal and Caste Populations
2003
[22]
논문
Mitochondrial lineage M1 traces an early human backflow to Africa
2007
[23]
논문
Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes
2005-05-13
[24]
논문
Revealing the prehistoric settlement of Australia by Y chromosome and mtDNA analysis
2007
[25]
논문
Mitochondrial DNA structure in the Arabian Peninsula
2008
[26]
논문
Mitochondrial DNA haplogroup K is associated with a lower risk of Parkinson's disease in Italians
2005
[27]
서적
The Evolution and History of Human Populations in South Asia
2007
[28]
논문
Multiplexed SNP Typing of Ancient DNA Clarifies the Origin of Andaman mtDNA Haplogroups amongst South Asian Tribal Populations
2006-12-20
[29]
논문
The 200,000-Year Evolution of Homo sapiens sapiens Language and Myth Families based on the mtDNA Phylotree, Fossil mtDNA and Archaeology: A Thought Experiment
2014
[30]
논문
A Mitochondrial Stratigraphy for Island Southeast Asia
2007
[31]
논문
A mitochondrial stratigraphy for island southeast Asia
2007
[32]
논문
Multiplex amplified product-length polymorphism analysis of 36 mitochondrial single-nucleotide polymorphisms for haplogrouping of East Asian populations
2005
[33]
논문
Utility of haplogroup determination for forensic mtDNA analysis in the Japanese population
2007
[34]
논문
Major Population Expansion of East Asians Began before Neolithic Time: Evidence of mtDNA Genomes
2011
[35]
논문
Most of the extant mtDNA boundaries in south and southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans
2004
[36]
논문
Mitochondrial DNA Haplogroup Analysis Reveals no Association between the Common Genetic Lineages and Prostate Cancer in the Korean Population
[37]
논문
The Peopling of Korea Revealed by Analyses of Mitochondrial DNA and Y-Chromosomal Markers
[38]
논문
Mitochondrial DNA haplogroups and homogeneity in the Korean population
[39]
논문
Tracing the Austronesian Footprint in Mainland Southeast Asia: A Perspective from Mitochondrial DNA
[40]
논문
Southeast Asian diversity: first insights into the complex mtDNA structure of Laos
[41]
논문
The Impact of the Austronesian Expansion: Evidence from mtDNA and Y Chromosome Diversity in the Admiralty Islands of Melanesia
2008-07-01
[42]
논문
Revealing the prehistoric settlement of Australia by Y chromosome and mtDNA analysis
[43]
논문
Genetic structure among Fijian island populations
[44]
논문
Analyses of genetic data within an interdisciplinary framework to investigate recent human evolutionary history and complex disease
https://ufdc.ufl.edu[...]
2010-01-01
[45]
학회
MtDNA variation in North, East, and Central African populations gives clues to a possible back-migration from the Middle East
2005-04-01
[46]
논문
Linking the sub-Saharan and West Eurasian gene pools: maternal and paternal heritage of the Tuareg nomads from the African Sahel
2010-03-17
[47]
논문
mtDNA from the Early Bronze Age to the Roman Period Suggests a Genetic Link between the Indian Subcontinent and Mesopotamian Cradle of Civilization
2013-01-01
[48]
논문
Pleistocene Mitochondrial Genomes Suggest a Single Major Dispersal of Non-Africans and a Late Glacial Population Turnover in Europe
2016-03-21
[49]
논문
Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations
2018-05-04
[50]
bioRxiv
Archaeogenetics of Late Iron Age Çemialo Sırtı, Batman: Investigating maternal genetic continuity in North Mesopotamia since the Neolithic
[51]
논문
Ancient Egyptian mummy genomes suggest an increase of Sub-Saharan African ancestry in post-Roman periods
2017-01-01
[52]
bioRxiv
Ancient genomes from North Africa evidence prehistoric migrations to the Maghreb from both the Levant and Europe
[53]
논문
The kinship of two 12th Dynasty mummies revealed by ancient DNA sequencing
https://pure.manches[...]
2018-02-01
[54]
논문
Large-Scale mtDNA Screening Reveals a Surprising Matrilineal Complexity in East Asia and Its Implications to the Peopling of the Region
2011-01-01
[55]
논문
Evolutionary History of Continental Southeast Asians: 'Early Train' Hypothesis Based on Genetic Analysis of Mitochondrial and Autosomal DNA Data
[56]
웹사이트
YFull MTree 1.01.14609
https://www.yfull.co[...]
2019-08-21
[57]
논문
Validation of microarray-based resequencing of 93 worldwide mitochondrial genomes
2009-01-01
[58]
논문
Human paternal and maternal demographic histories: insights from high-resolution Y chromosome and mtDNA sequences
[59]
논문
Complete Mitochondrial DNA Diversity in Iranians
[60]
논문
Mitochondrial and Y-chromosome diversity of the Tharus (Nepal): a reservoir of genetic variation
[61]
논문
The mitochondrial genomes of two Pre-historic Hunter Gatherers in Sri Lanka
2023-02-01
[62]
논문
Updating Phylogeny of Mitochondrial DNA Macrohaplogroup M in India: Dispersal of Modern Human in South Asian Corridor
[63]
논문
Mitochondrial DNA Variability in Slovaks, with Application to the Roma Origin
[64]
논문
New insights from Thailand into the maternal genetic history of Mainland Southeast Asia
[65]
논문
Tracing the legacy of the early Hainan Islanders - a perspective from mitochondrial DNA
[66]
논문
Mitochondrial Genome Diversity in the Tubalar, Even, and Ulchi: Contribution to Prehistory of Native Siberians and Their Affinities to Native Americans
[67]
논문
Autosomal and uniparental portraits of the native populations of Sakha (Yakutia): implications for the peopling of Northeast Eurasia
[68]
논문
Investigating the Prehistory of Tungusic Peoples of Siberia and the Amur-Ussuri Region with Complete mtDNA Genome Sequences and Y-chromosomal Markers
[69]
논문
Phylogeographic Analysis of Mitochondrial DNA in Northern Asian Populations
[70]
논문
Mitochondrial Genome Variation in Eastern Asia and the Peopling of Japan
2004-10-01
[71]
논문
Fine-Scale Population Admixture Landscape of Tai–Kadai-Speaking Maonan in Southwest China Inferred From Genome-Wide SNP Data
2022-02-17
[72]
논문
The matrilineal ancestry of Nepali populations
2022-10-15
[73]
논문
A Late Pleistocene human genome from Southwest China
2022-07
[74]
논문
(Title missing)
2024-07
[75]
논문
Diverse origin of mitochondrial lineages in Iron Age Black Sea Scythians
2017
[76]
논문
Mitochondrial genomes uncover the maternal history of the Pamir populations
2018-01
[77]
논문
Cultural variation impacts paternal and maternal genetic lineages of the Hmong-Mien and Sino-Tibetan groups from Thailand
2020-11
[78]
논문
Early Austronesians: into and out of Taiwan
2014
[79]
논문
East Asian mtDNA haplogroup determination in Koreans: Haplogroup-level coding region SNP analysis and subhaplogroup-level control region sequence analysis
2006-11
[80]
논문
New insights into the fine-scale history of western–eastern admixture of the northwestern Chinese population in the Hexi Corridor via genome-wide genetic legacy
2021-05
[81]
논문
Mitochondrial DNA Sequence Variation and Haplogroup Distribution in Chinese Patients with LHON and m.14484T>C
2010-10-18
[82]
논문
Mitochondrial DNA Haplogroups M7b1′2 and M8a Affect Clinical Expression of Leber Hereditary Optic Neuropathy in Chinese Families with the m.11778G→A Mutation
2008-12
[83]
논문
Ancestral Origins and Genetic History of Tibetan Highlanders
2016-09
[84]
논문
MtDNA analysis reveals enriched pathogenic mutations in Tibetan highlanders
2016-08-08
[85]
논문
Large-scale mitochondrial DNA analysis in Southeast Asia reveals evolutionary effects of cultural isolation in the multi-ethnic population of Myanmar
2014-12
[86]
논문
Rate variation between mitochondrial domains and adaptive evolution in humans
2007-10
[87]
논문
Full mtGenome reference data: Development and characterization of 588 forensic-quality haplotypes representing three U.S. populations
2015-01
[88]
논문
Phylogeny of East Asian Mitochondrial DNA Lineages Inferred from Complete Sequences
2003-09
[89]
논문
The MT-ND1 and MT-ND5 genes are mutational hotspots for Chinese families with clinical features of LHON but lacking the three primary mutations
2010-08
[90]
논문
New insights from Thailand into the maternal genetic history of Mainland Southeast Asia
2018-06
[91]
논문
Large-Scale mtDNA Screening Reveals a Surprising Matrilineal Complexity in East Asia and Its Implications to the Peopling of the Region
2011-01
[92]
논문
Mitogenomic diversity in Russians and Poles
2017-09
[93]
논문
Sex-specific genetic diversity is shaped by cultural factors in Inner Asian human populations
2017-04
[94]
논문
Strikingly different penetrance of LHON in two Chinese families with primary mutation G11778A is independent of mtDNA haplogroup background and secondary mutation G13708A
2008-08
[95]
논문
East Eurasian ancestry in the middle of Europe: genetic footprints of Steppe nomads in the genomes of Belarusian Lipka Tatars
2016-07-25
[96]
논문
Mitogenomic diversity and differentiation of the Buryats
2018-01
[97]
논문
The History of Slavs Inferred from Complete Mitochondrial Genome Sequences
2013-01-14
[98]
논문
Investigating Holocene human population history in North Asia using ancient mitogenomes
2018-06-12
[99]
논문
Mitogenomic Diversity in Tatars from the Volga-Ural Region of Russia
2010-10
[100]
논문
Deciphering the Signature of Selective Constraints on Cancerous Mitochondrial Genome
2012-04
[101]
논문
Mitochondrial DNA association study of type 2 diabetes with or without ischemic stroke in Taiwan
2014-12
[102]
논문
Mitochondrial Haplogroups N9 and G Are Associated with Metabolic Syndrome Among Human Immunodeficiency Virus-Infected Patients in China
2019-06
[103]
논문
Mitochondrial genome diversity on the Central Siberian Plateau with particular reference to the prehistory of northernmost Eurasia
2021-01-28
[104]
논문
Uncovering the Profile of Somatic mtDNA Mutations in Chinese Colorectal Cancer Patients
2011-06-28
[105]
논문
Co-occurrence of A1555G and G11778A in a Chinese family with high penetrance of Leber's hereditary optic neuropathy
2008-11
[106]
논문
[141]
웹사이트
M77 MTree
https://yfull.com/mt[...]
[142]
웹사이트
M62'68 MTree
https://yfull.com/mt[...]
[143]
웹사이트
M68 MTree
https://yfull.com/mt[...]
[144]
논문
Human population history at the crossroads of East and Southeast Asia since 11,000 years ago
2021-07-00
[145]
논문
Contrasting maternal and paternal genetic variation of hunter-gatherer groups in Thailand
2018-01-24
[146]
웹사이트
M73'79 MTree
https://yfull.com/mt[...]
[147]
웹사이트
M73 MTree
https://yfull.com/mt[...]
[148]
웹사이트
M79 MTree
https://yfull.com/mt[...]
[149]
논문
Admixture, migrations, and dispersals in Central Asia: evidence from maternal DNA lineages
2004-06-00
[150]
논문
Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation
2009-00-00
[151]
잡지
The Moment Magazine Great DNA Experiment
https://momentmag.co[...]
2012-09-00
[152]
잡지
The Moment Magazine Great DNA Experiment
https://momentmag.co[...]
2012-09-00
[153]
논문
Most of the extant mtDNA boundaries in South and Southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans
2004-00-00
[154]
서적
The evolution and history of human populations in South Asia
https://books.google[...]
Springer
[155]
논문
Multiplexed SNP Typing of Ancient DNA Clarifies the Origin of Andaman mtDNA Haplogroups amongst South Asian Tribal Populations
2006-12-20
[156]
논문
Phylogeny and antiquity of M macrohaplogroup inferred from complete mt DNA sequence of Indian specific lineages
2005-00-00
[157]
논문
The Peopling of Korea Revealed by Analyses of Mitochondrial DNA and Y-Chromosomal Markers
2009-00-00
[158]
논문
Philippine Mitochondrial DNA Diversity: A Populated Viaduct between Taiwan and Indonesia?
2010-01-00
[159]
웹인용
MtDNA variation in North, East, and Central African populations gives clues to a possible back-migration from the Middle East
http://konig.la.utk.[...]
American Association of Physical Anthropologists
2016-04-13
[160]
논문
Linking the sub-Saharan and West Eurasian gene pools: maternal and paternal heritage of the Tuareg nomads from the African Sahel
2010-03-17
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com